1.Deisseroth K, G Feng, AK Majewska, G Miesenbock, A Ting and MJ Schnitzer. (2006). Next-generation optical technologies for illuminating genetically targeted brain circuits. J Neurosci 26:10380-6.
2.Li C, W Sun, C Gu, Z Yang, N Quan, J Yang, Z Shi, L Yu and H Ma. (2018). Targeting ALDH2 for Therapeutic Interventions in Chronic Pain-Related Myocardial Ischemic Susceptibility. Theranostics 8:1027-1041.
3.Yuan Y, Y Zheng, X Zhang, Y Chen, X Wu, J Wu, Z Shen, L Jiang, L Wang, W Yang, J Luo, Z Qin, W Hu and Z Chen. (2017). BNIP3L/NIX-mediated mitophagy protects against ischemic brain injury independent of PARK2. Autophagy 13:1754-1766.
4.Li S, X Dou, H Ning, Q Song, W Wei, X Zhang, C Shen, J Li, C Sun and Z Song. (2017). Sirtuin 3 acts as a negative regulator of autophagy dictating hepatocyte susceptibility to lipotoxicity. Hepatology 66:936-952.
5.Li L, B Li, M Li, C Niu, G Wang, T Li, E Krol, W Jin and JR Speakman. (2017). Brown adipocytes can display a mammary basal myoepithelial cell phenotype in vivo. Mol Metab 6:1198-1211.
6.Feng D, B Wang, L Wang, N Abraham, K Tao, L Huang, W Shi, Y Dong and Y Qu. (2017). Pre-ischemia melatonin treatment alleviated acute neuronal injury after ischemic stroke by inhibiting endoplasmic reticulum stress-dependent autophagy via PERK and IRE1 signalings. J Pineal Res 62.
7.Du X, H Hao, Y Yang, S Huang, C Wang, S Gigout, R Ramli, X Li, E Jaworska, I Edwards, J Deuchars, Y Yanagawa, J Qi, B Guan, DB Jaffe, H Zhang and N Gamper. (2017). Local GABAergic signaling within sensory ganglia controls peripheral nociceptive transmission. J Clin Invest 127:1741-1756.
8.Yang H, J Yang, W Xi, S Hao, B Luo, X He, L Zhu, H Lou, YQ Yu, F Xu, S Duan and H Wang. (2016). Laterodorsal tegmentum interneuron subtypes oppositely regulate olfactory cue-induced innate fear. Nat Neurosci 19:283-9.
9.Wu X, X Wu, Y Ma, F Shao, Y Tan, T Tan, L Gu, Y Zhou, B Sun, Y Sun, X Wu and Q Xu. (2016). CUG-binding protein 1 regulates HSC activation and liver fibrogenesis. Nat Commun 7:13498.
10.Wei Y, Y Chen, Y Qiu, H Zhao, G Liu, Y Zhang, Q Meng, G Wu, Y Chen, X Cai, H Wang, H Ying, B Zhou, M Liu, D Li and Q Ding. (2016). Prevention of Muscle Wasting by CRISPR/Cas9-mediated Disruption of Myostatin In vivo. Mol Ther 24:1889-1891.
11.Zhang X, Y Yuan, L Jiang, J Zhang, J Gao, Z Shen, Y Zheng, T Deng, H Yan, W Li, WW Hou, J Lu, Y Shen, H Dai, WW Hu, Z Zhang and Z Chen. (2014). Endoplasmic reticulum stress induced by tunicamycin and thapsigargin protects against transient ischemic brain injury: Involvement of PARK2-dependent mitophagy. Autophagy 10:1801-13.





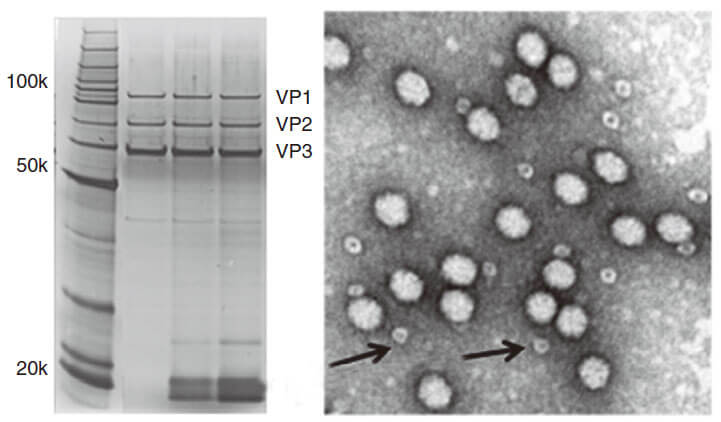
 Lentivirus User Manual.
1. For detailed protocol about how to package and purify Adv, please see the
Lentivirus User Manual.
1. For detailed protocol about how to package and purify Adv, please see the 